Introduction and Scope
Nontuberculous mycobacteria (NTM) have long been regarded as environmental opportunists rather than classical pathogens. However, a series of systematic reviews and surveillance analyses published up to 2025 emphasize their growing clinical and veterinary impact, as well as the underappreciated zoonotic potential that has been largely overlooked in earlier literature (1–3). More than 200 species of NTM have now been described, spanning rapid and slow growers; however, only approximately 20 species have credible evidence supporting zoonotic transmission or animal–human linkage. These include Mycobacterium marinum in aquaculture-related cutaneous infections and Mycobacterium avium complex (MAC) in avian and porcine reservoirs (4–6).
Zoonotic NTM are particularly relevant at the human–animal–environment interface. Outbreak investigations implicate aquaculture workers, veterinarians, dairy handlers, and laboratory staff as occupationally at risk, with infection routes often mediated by water systems and biofilms (4,5). Climate warming is now recognized as an important ecological driver: elevated water temperatures and altered pH promote NTM persistence and proliferation in plumbing networks, cooling devices, and natural reservoirs. These changes expand the zoonotic interface by increasing the bacterial load in aquatic systems that intersect with livestock, aquaculture, and human use (7–10).
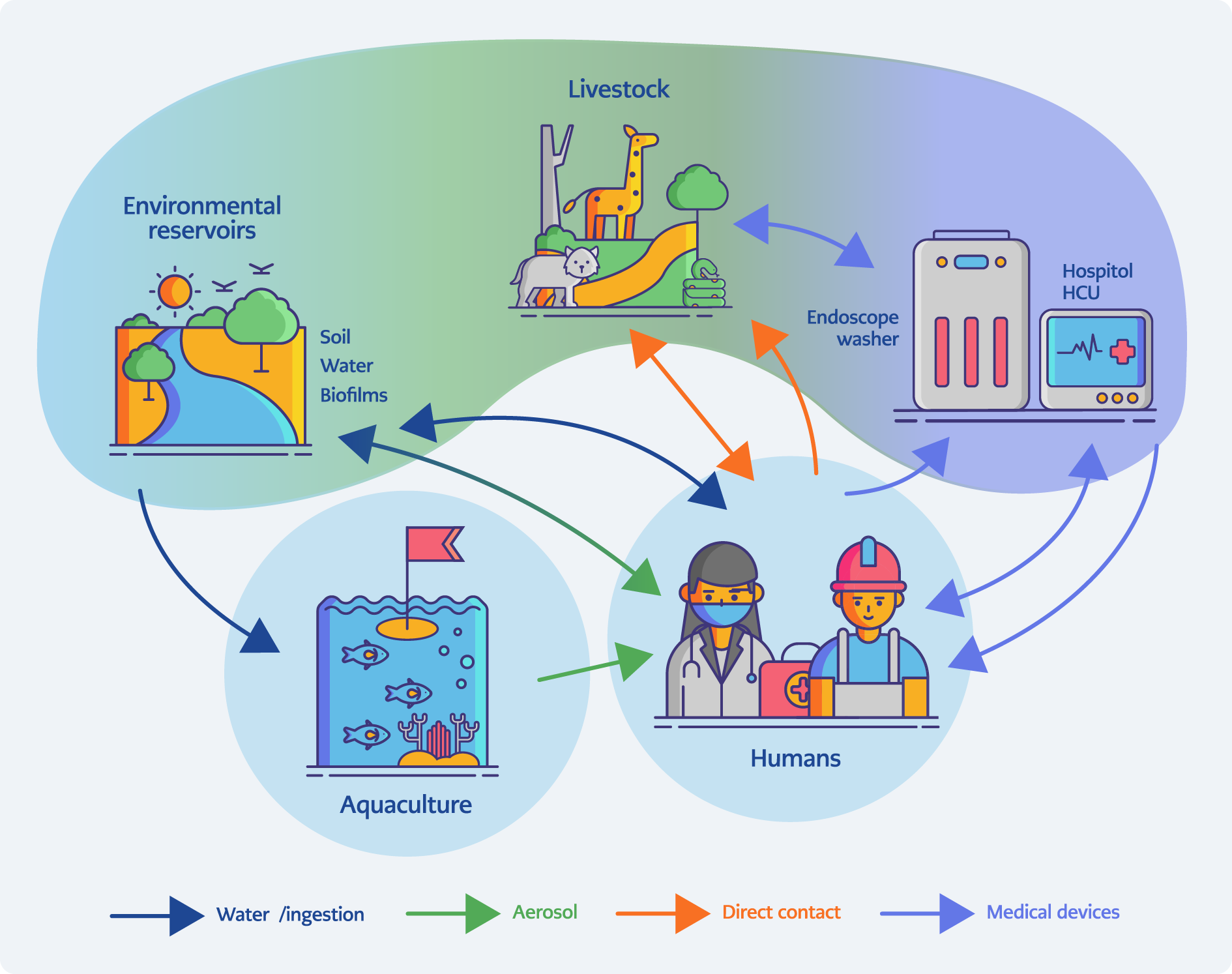
Figure 1. One Health transmission network for zoonotic nontuberculous mycobacteria (NTM). Conceptual framework illustrating how environmental reservoirs (soil, water, biofilms), aquaculture, livestock and wildlife, engineered water systems (e.g., heater-cooler units and endoscope washers), and human occupational or clinical settings are interconnected. Arrows indicate major exposure pathways including aerosolization, ingestion, direct contact, and device-mediated transmission, while color bands highlight dominant ecological routes for exemplar species (e.g., aquatic M. marinum and device-associated M. chimaera). [Graphically prepared with the assistance of ChatGPT (OpenAI, San Francisco, USA); all data interpretation and final figure content were verified by the authors.]
The scope of this review is to critically appraise the current understanding of zoonotic NTM transmission, clinical and laboratory diagnosis, and methodological advances in detection, while situating the discussion within a One Health and climate-sensitive framework (11,12). We highlight both established zoonotic NTM species and those of emerging concern, emphasizing the implications for diagnostics, surveillance, and antimicrobial resistance (AMR) monitoring in an era of global environmental change. Figure 1 summarizes the One Health framework linking environmental, animal, engineered, and human reservoirs through the major transmission pathways that drive zoonotic NTM exposure.
In this review, we use “true zoonosis” to denote events where robust epidemiologic and genomic evidence demonstrates direct or indirect transmission of a pathogen from an animal host to a human (i.e., animal-to-human transmission supported by temporally linked isolates and high-resolution genomic concordance). We use “common-source exposure” to denote scenarios in which humans and animals are independently exposed to the same contaminated environmental reservoir (e.g., a shared water system), without evidence of direct animal-to-human transmission. Where evidence is insufficient to distinguish between these possibilities, we adopt the more neutral term “animal-linked” or “animal-associated” exposure. This distinction is critical for surveillance, outbreak attribution, and control measures, and is applied consistently throughout the manuscript.
Materials and Methods
We performed a targeted narrative review using PubMed, Scopus, and Web of Science (search window: January 1, 2018, to September 30, 2025). Primary search terms included “nontuberculous mycobacteria”, “NTM”, “zoonotic mycobacteria”, “Mycobacterium chimaera”, “Mycobacterium marinum”, “heater–cooler unit”, “aquaculture mycobacteria”, “metagenomics”, “whole-genome sequencing”, “PMA-ddPCR”, and “antimicrobial resistance”. We supplemented database searches with focused queries of device-related advisories from the U.S. Food and Drug Administration (FDA) and Centers for Disease Control and Prevention (CDC) as One Health policy documents, and reviewed reference lists of key articles.
We included original research articles, outbreak reports, surveillance studies, systematic reviews, methodological papers, and policy statements that directly addressed zoonotic or animal-linked NTM, diagnostic methodologies, biosafety, or One Health surveillance. Exclusion criteria were conference abstracts without full text, non-English articles without an English abstract, and papers that did not present primary data or substantive methodological detail. This review emphasizes species-specific and methodological literature relevant to zoonotic transmission and laboratory detection.
Key Genomic Reclassifications and Taxonomic Updates
Nontuberculous mycobacteria are taxonomically diverse. Below, we summarize recent genomics-driven reclassifications with clinical and zoonotic relevance. Traditional classification relied on phenotypic traits such as growth rate, pigment production, and biochemical profiles, but these approaches have proven insufficient for delineating closely related taxa. The advent of multilocus sequence typing and, more decisively, whole-genome sequencing (WGS), has restructured the taxonomy of several clinically important groups (8–10).
For instance, the Mycobacterium abscessus complex—long considered a single entity—has been subdivided into M. abscessus subsp. abscessus, M. abscessus subsp. massiliense, and M. abscessus subsp. bolletii (13–15). These reclassifications are clinically meaningful because macrolide resistance hinges on the functionality of the erm(41) gene, which differs between subspecies and directly influences therapeutic outcomes. Similar genomic clarifications have emerged in the MAC, including the separation of M. avium subsp. avium, M. avium subsp. hominissuis, and others (5).

Table 1. Selected NTM reclassifications by whole-genome sequencing.
As sequencing capacity expands, WGS-based phylogenomics is replacing phenotypic descriptors as the taxonomic backbone for NTM, providing higher resolution for outbreak investigation, zoonotic attribution, and resistance gene mapping (8–10). For clinicians and veterinarians, these reclassifications underscore the need to integrate genomic data into both diagnostic workflows and antimicrobial stewardship strategies (Table 1) (16–18).
Epidemiology and Reservoirs
The epidemiology of zoonotic NTM is heterogeneous, reflecting the broad ecological distribution of these organisms in soil, water, animals, and engineered environments. Historically viewed as sporadic opportunists, NTM are now increasingly implicated in systematic outbreaks and occupational exposures, with recent data from 2023–2025 underscoring their relevance at the human–animal–environment interface (1,3).
Domestic and Food-Animal Reservoirs
Several zoonotic NTM species colonize livestock and poultry, creating both direct and indirect pathways for human exposure. Recent surveillance in China (2023–2024) demonstrated M. abscessus contamination within cattle farms and slaughterhouse effluents, highlighting the zoonotic risks embedded in food-production chains (19). M. avium subsp. hominissuis remains widely detected in swine herds and dairy cattle, where it serves as a source of zoonotic infections for farmers, veterinarians, and abattoir workers (6).
Camels and Arid-Region Reservoirs
Emerging reports from the Middle East and sub-Saharan Africa point to camels as reservoirs of multiple NTM species. In these arid ecosystems, Mycobacterium fortuitum and Mycobacterium intracellulare have been isolated from nasal swabs and milk, suggesting potential zoonotic transmission routes through both occupational contact and dairy consumption (20). The camel-NTM interface warrants close monitoring, as these animals sustain livelihoods across fragile, climate-stressed regions where diagnostic infrastructure is often limited.
Wildlife Reservoirs
Wildlife contributes to the persistence and transmission of certain zoonotic NTM. A well-studied example is the role of possums in the ecology of Mycobacterium ulcerans in Australia, where fecal shedding and environmental contamination sustain transmission cycles (21,22). Similar wildlife-associated reservoirs are increasingly recognized in other regions, though their zoonotic significance remains incompletely mapped.
Hospital Water Systems and Engineered Environments
Beyond natural reservoirs, hospital and industrial water systems serve as amplifiers of the NTM burden. Biofilm formation within plumbing networks, endoscope washers, and heater-cooler units (HCUs) creates a resilient reservoir for persistent contamination. The global outbreaks of Mycobacterium chimaera linked to HCUs illustrate how engineered water systems, once colonized, can drive nosocomial epidemics with devastating consequences (5). Such events blur the distinction between environmental and healthcare-associated reservoirs.
Evidence from India confirms that NTM are not only opportunistic clinical pathogens but are also entrenched in community environments. In Wardha district (central India), a two-year environmental surveillance study tested 1398 soil and water samples and recovered NTM from 12.2% of samples, with 63% classified as potentially pathogenic mycobacteria. Isolation was highest from soil used for community gatherings (42.8%), residential premises (27.7%), and well surroundings (26.0%), while drinking water wells (15.4%), treated storage tanks (6.9%), and household receptacles (6.3%) were the most contaminated potable sources. M. fortuitum, Mycobacterium gordonae and MAC were dominant species, followed by Mycobacterium flavescens, Mycobacterium scrofulaceum, Mycobacterium simiae and M. marinum. Seasonal analysis showed peak NTM isolation during the monsoon (20.3%), falling to 5.8% in summer, demonstrating strong climate sensitivity of environmental NTM exposure (23).
One Health and Policy Perspectives
Epidemiological data on zoonotic NTM fit squarely into a One Health paradigm. The United States National One Health Framework for 2025–2029 prioritizes zoonotic pathogen surveillance, explicitly including atypical and emerging mycobacteria within its scope. This policy alignment reflects the growing recognition that NTM outbreaks in humans are rarely explainable without considering parallel animal and environmental data (11,12). Integrating these surveillance layers offers not only early outbreak detection but also insights into the ecological and climatic drivers that expand NTM reservoirs.
Region-Specific Burden and Trends
In the United States, population-based analyses show sustained increases in NTM pulmonary disease (NTM-PD). Among adults aged ≥65 years (2008–2015), incidence rose from 12.70 to 18.37 per 100,000 person-years and prevalence increased from 30.27 to 47.48 per 100,000 persons (24). In a large United States Medicare cohort (2010–2019), the average annual incidence of NTM-PD was 20.1 per 100,000, with the highest rates in the southern United States and increasing annual percent change across regions (notably Northeast and Midwest) (25).
In Europe, nationwide registry data from Denmark (1991–2022) similarly demonstrate increasing pulmonary NTM incidence, with marked age and sex gradients (higher rates in older females) and geographic heterogeneity across municipalities (26). In East Asia, South Korea reported a sharp rise in NTM disease burden over the last decade: annual prevalence increased from 11.4 to 56.7 per 100,000 population between 2010 and 2021, and among persons aged ≥65 years, prevalence rose from 41.9 to 163.1 per 100,000 (27). In Taiwan, a population-based retrospective cohort showed age-adjusted incidence increasing from 5.3 to 14.8 per 100,000 per year between 2005 and 2013 (28). In India, robust national incidence estimates remain limited because NTM is not uniformly captured in routine surveillance; however, a synthesis focused on Indian settings reported that the prevalence of NTM-PD among presumptive tuberculosis patients was 1.1% (395/34,829), with pulmonary isolates most frequently MAC (19%) followed by Mycobacterium chelonae (10%), M. fortuitum (9.8%), and M. abscessus (8.8%) (29). A tertiary-care study from North India also characterized the burden and species diversity among clinical isolates, supporting substantial geographic variation within the country (30).
Clinical and Veterinary Syndromes
Zoonotic NTM manifest across a wide clinical spectrum in humans, ranging from self-limiting cutaneous infections to severe, chronic pulmonary disease. The clinical burden has shifted notably in recent years. While MAC historically dominated pulmonary NTM disease, registry data published in 2025 reveal an emerging dominance of M. abscessus in chronic lung infections, particularly among individuals with structural lung disease, cystic fibrosis, and post-tuberculosis sequelae (1,15,31). This shift is clinically significant: M. abscessus infections are notoriously recalcitrant to therapy, with intrinsic macrolide resistance linked to the erm(41) gene expression, complicating treatment regimens and driving poor outcomes.
Cutaneous and soft-tissue infections remain strongly associated with zoonotic and environmental exposures. M. marinum is classically linked to fish handlers and aquaculture workers (“fish tank granuloma”), while M. fortuitum and M. chelonae appear in postsurgical and posttraumatic wound infections where contaminated water or animal contact is implicated (4,6). Disseminated infections, though uncommon, occur in immunocompromised hosts, underscoring the opportunistic potential of zoonotic NTM once host defenses are impaired (6).
In the veterinary domain, NTM cause both sporadic disease and subclinical colonization across diverse species. Swine, cattle, poultry, and camels have all been reported to harbor M. avium complex, M. intracellulare, and rapid growers such as M. fortuitum (19,20). Companion animals, including dogs and cats, also occasionally present with cutaneous or disseminated NTM infections, serving as potential reservoirs. Recent reports highlight the AMR dimension in veterinary isolates: M. abscessus and M. fortuitum strains recovered from domestic and wild animals exhibit resistance profiles overlapping those of human clinical isolates, raising concerns about zoonotic transmission of drug-resistant strains (32,33).
Evidence from animal surveillance and clinical veterinary microbiology indicates that both rapidly growing and slow-growing NTM recovered from animals can carry clinically relevant resistance patterns. In a Croatian national surveillance dataset of 87 rapidly growing mycobacteria (RGM) isolates from domestic and wild animals (2012–2015) tested against 15 antimicrobials, the highest resistance frequencies were reported to cephalosporins (>50%), followed by amoxicillin–clavulanate (31.0%), clarithromycin (23.0%), tobramycin (14.9%), and doxycycline (10.3%), while resistance to imipenem, ciprofloxacin, minocycline, and linezolid was <7%; importantly, the authors reported universal susceptibility to amikacin and moxifloxacin in their tested isolate set (33). These findings highlight that animal-associated RGM can include macrolide-nonsusceptible isolates (a critical clinical issue for zoonotic interface pathogens), while also suggesting that some parenteral backbones (e.g., amikacin) may retain activity in certain settings.
Complementing surveillance data, clinical veterinary isolates from companion animals show substantial species- and drug-specific variability. In Taiwan, 44 RGM isolates from dogs and cats (2018–2021) were dominated by M. fortuitum complex (20 isolates) and M. abscessus complex (20 isolates); >80% of isolates were susceptible to linezolid and amikacin, whereas M. abscessus complex isolates showed multidrug resistance patterns (reported as resistance to ≥3 major antibiotic groups in the study) with approximately 75% susceptible to clarithromycin, while only approximately 35% of M. fortuitum complex isolates were susceptible to clarithromycin, consistent with the clinical challenge of inducible or species-dependent macrolide nonsusceptibility in animal-associated RGM (34).
Resistance is not confined to RGM. A Croatian analysis of 44 slow-growing NTM isolates from domestic and wild animals tested against 13 antimicrobials found high resistance proportions to moxifloxacin (77.3%), doxycycline (76.9%), and rifampicin (76.9%), with additional resistance to ciprofloxacin (65.4%), trimethoprim-sulfamethoxazole (65.4%), and linezolid (61.4%); resistance to rifabutin (7.7%) and amikacin (6.8%) was low, and no isolates were resistant to clarithromycin in that dataset (35). Together, these animal data support a One Health rationale for integrating veterinary and wildlife isolates into AMR surveillance—particularly at human–animal–environment interfaces—while also emphasizing that antimicrobial susceptibility testing (AST) interpretation must remain species- and method-specific and may not generalize across regions or host populations.
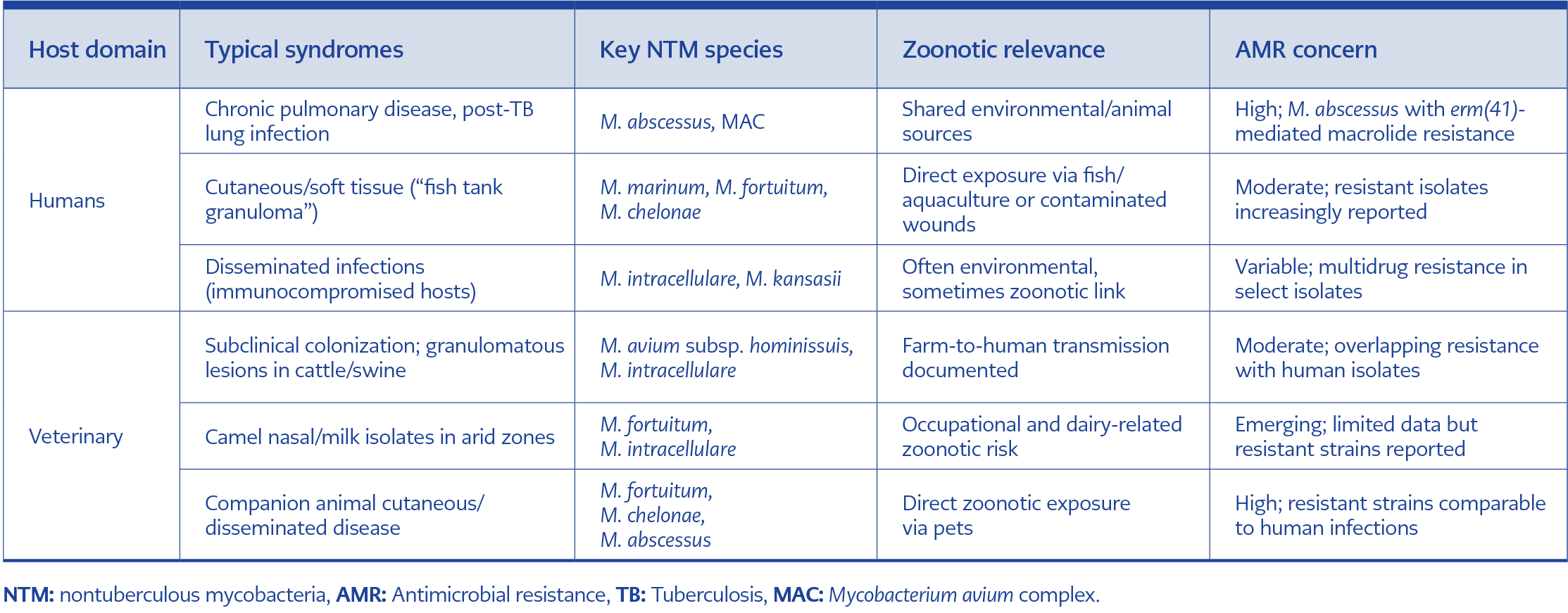
Table 2. Comparative clinical and veterinary syndromes of zoonotic NTM.
Veterinary and wildlife NTM isolates can show meaningful macrolide and multidrug resistance patterns, so clinically significant animal-associated isolates should undergo standardized broth microdilution AST (or referral testing) to inform zoonotic-risk interpretation and cross-sector antimicrobial stewardship. The overlap between human and animal disease emphasizes a continuum of clinical syndromes (Table 2), rather than discrete reservoirs. For clinicians and veterinarians alike, this convergence demands coordinated diagnostic vigilance and AMR monitoring to capture the true scope of zoonotic NTM.
Transmission Pathways
The routes of transmission for zoonotic NTM are diverse but increasingly understood through molecular epidemiology and experimental studies. Recent work confirms that aerosol exposure remains the primary transmission route for both environmental and zoonotic NTM. Inhalation of bioaerosols generated from contaminated soil, animal enclosures, farms, and slaughterhouse effluents has been documented as a significant driver of pulmonary disease in agricultural workers. Such airborne dissemination is particularly relevant for rapid growers like M. abscessus and M. fortuitum, which thrive in moist organic matter and water systems capable of aerosolization (19,36).
Ingestion and wound inoculation form important secondary routes. Aquaculture-linked infections by M. marinum and cutaneous outbreaks of M. chelonae often trace back to minor trauma in contaminated aquatic environments. Ingestion of raw or under-processed animal products has been associated with sporadic NTM detections in humans; however, the evidence is primarily from case clusters and molecular matching rather than controlled epidemiologic studies and should therefore be interpreted cautiously (33).
Community-level exposure through soil and drinking water is a major but under-recognized transmission interface in tuberculosis-endemic regions. In rural central India, Wardha district environmental surveillance demonstrated that potable water sources (wells, tanks, household containers, taps) and communal soils were heavily contaminated with potentially pathogenic NTM, including MAC and M. fortuitum. These reservoirs coincide spatially with human congregation sites and domestic water use, creating continuous opportunities for aerosol, ingestion, and cutaneous exposure—particularly during the rainy season when NTM recovery peaked (23).
At the cellular level, NTM demonstrate sophisticated pathogenesis mechanisms that facilitate persistence, chief among them the ability to evade host autophagy pathways. Experimental models have shown that species such as M. abscessus and MAC disrupt phagosome–lysosome fusion, thereby avoiding intracellular killing and establishing chronic infection. This autophagy evasion contributes to their ability to colonize immunocompromised hosts and survive in hostile environments (37).
Host susceptibility factors increasingly modify transmission risk. The expanding global use of biologic immunomodulators, such as tumor necrosis factor (TNF) inhibitors and interleukin-targeted therapies, has emerged as a key driver of opportunistic NTM disease. These drugs impair granuloma formation and intracellular killing, effectively lowering the host’s threshold for NTM infection following environmental or zoonotic exposure (38).
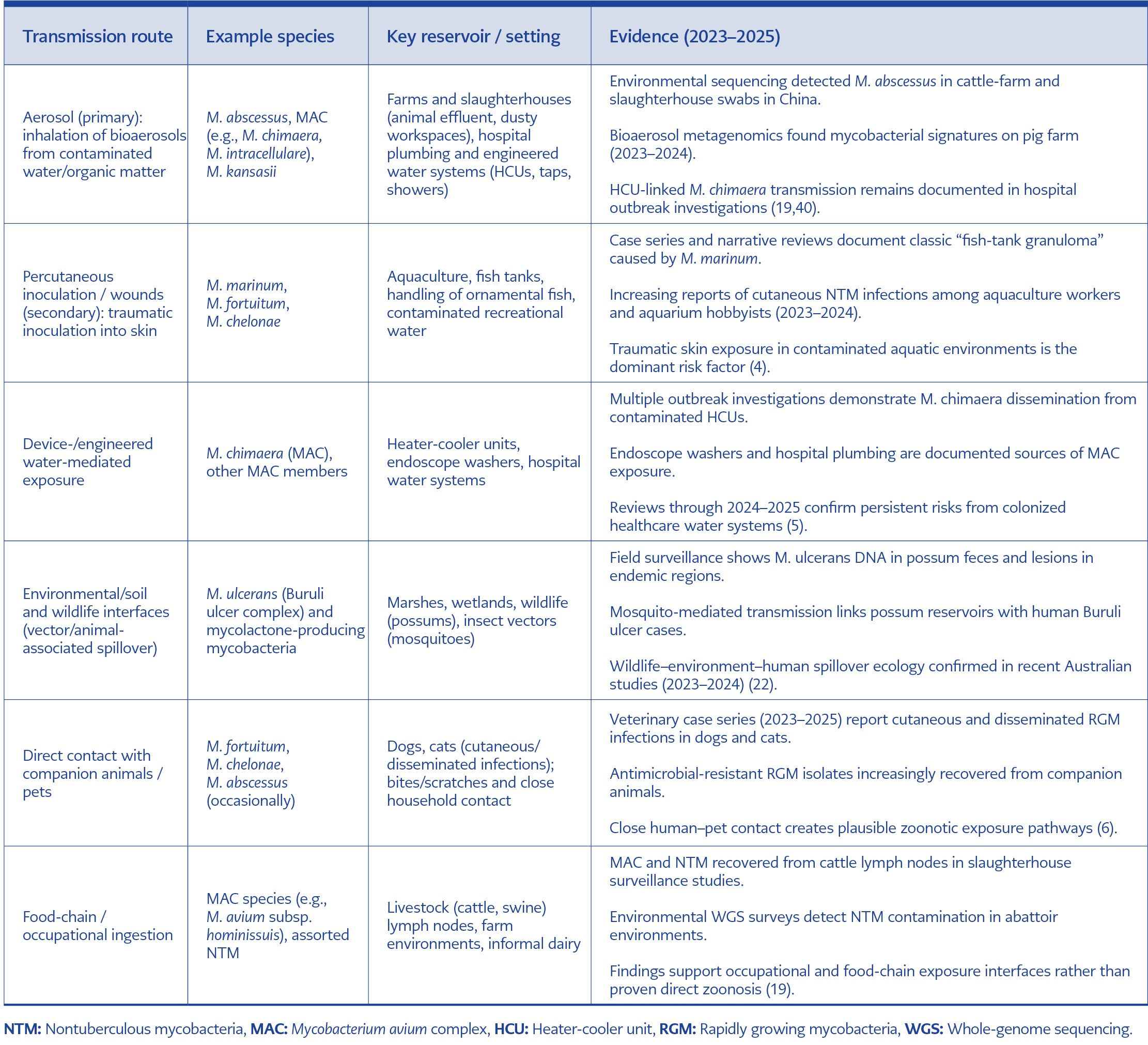
Table 3. Evidence-based transmission route.
Together, these pathways emphasize the multimodal transmission ecology of zoonotic NTM, where aerosol predominates, but ingestion, traumatic inoculation, and host susceptibility modifiers critically shape clinical outcomes. Recognizing the relative importance of these routes (Table 3) is essential for designing occupational safety protocols, veterinary biosecurity measures, and One Health-informed interventions (39).
Laboratory Diagnosis and Detection Methodologies
Specimen Strategy
Specimen selection for zoonotic NTM diagnosis requires careful alignment with the suspected clinical or veterinary syndrome. Respiratory samples (sputum, bronchoalveolar lavage) remain the cornerstone in pulmonary disease, while tissue biopsies, pus aspirates, and wound swabs are prioritized for cutaneous forms. In veterinary surveillance, lymph node aspirates and milk from mastitic animals are increasingly used to capture subclinical infections. Recent One Health-oriented field studies emphasize environmental sampling, including water biofilms, soil, and slaughterhouse aerosols, where the adoption of propidium monoazide (PMA)-treated assays coupled with droplet digital polymerase chain reaction (ddPCR) has allowed viability discrimination between live and dead organisms, mitigating false positives from DNA persistence in the environment (8,18,41).
Collect specimens that match the clinical or veterinary syndrome (respiratory, tissue, lymph node, milk, or water biofilm) and prioritize paired human–animal–environmental sampling whenever zoonotic transmission is suspected.
Culture Approaches
Decontamination protocols remain species-dependent. Standard N-acetyl-L-cysteine-sodium hydroxide (NALC-NaOH) treatment is effective for many slow growers, but modified oxalic acid protocols are recommended for samples with high contamination risk (e.g., animal feces). Optimal incubation temperatures vary: M. marinum and M. ulcerans grow best at 28–32°C, while most rapid growers, including M. abscessus, prefer 35–37°C. Solid media (Löwenstein–Jensen, Middlebrook 7H10/7H11) are complemented by liquid culture systems, such as mycobacterial growth indicator tube 960 (MGIT 960), which shorten detection times to 7–14 days for rapid growers. However, culture remains slow for pathogens like M. ulcerans, often requiring >8 weeks, highlighting the importance of molecular adjuncts (42).
Use temperature-appropriate solid and liquid mycobacterial culture (30–32°C for M. marinum/M. ulcerans, 35–37°C for most others) and avoid over-decontamination to maximize recovery of fastidious zoonotic NTM.
Identification (ID)
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) has matured into a frontline identification tool, though library gaps for uncommon zoonotic isolates persist. Updated reference databases in 2024–2025 now include spectra for veterinary isolates such as M. fortuitum and M. chelonae. Artificial intelligence (AI)-assisted classifiers can accelerate species-level calls from metagenomic data; recent pilot studies report 24–48 hour timeframes in specialized centers, but broader implementation remains limited. Whole-genome sequencing remains the gold standard for outbreak investigation and reclassification, exemplified by the ongoing refinement of subspecies differentiation within M. abscessus complex using erm(41)-based genotyping (8,39).
Apply MALDI-TOF or targeted sequencing for rapid species identification and reserve WGS for outbreak investigation, subspecies resolution, and zoonotic source attribution.
Antimicrobial Susceptibility Testing (AST)
The Clinical and Laboratory Standards Institute (CLSI) M24 (2023 update) and M62 (2024 guidance) documents remain the global standards for NTM AST. They emphasize the necessity of broth microdilution for reproducibility and the importance of detecting inducible macrolide resistance mediated by erm(41) expression in M. abscessus. Updated recommendations also clarify breakpoints for amikacin, clarithromycin, and tigecycline. Veterinary and zoonotic isolates frequently exhibit multidrug resistance, complicating cross-species management. Reports from 2024 to 2025 highlight rising linezolid resistance in M. fortuitum and aminoglycoside resistance in M. chelonae. In resource-limited settings, full CLSI-compliant panels are often not sustainable; targeted panels for macrolides, aminoglycosides, and fluoroquinolones have been proposed as pragmatic alternatives (16).
Perform CLSI-compliant broth microdilution testing with the mandatory macrolide (erm(41)) and amikacin for clinically significant isolates, and refer complex panels to reference laboratories when needed.
Molecular Detection
Molecular assays remain pivotal where culture is slow or unrewarding. Gene targets such as hsp65, rpoB, and the 16S–23S internal transcribed spacer (ITS) continue to underpin PCR assays for rapid speciation. Low-resource laboratories in low- and middle-income countries (LMICs) are adopting multiplex real-time PCR panels targeting common zoonotic NTM clusters (e.g., M. fortuitum group, M. chelonae–abscessus group). Meanwhile, ddPCR offers unparalleled sensitivity for environmental detection, especially when combined with PMA to exclude DNA from nonviable cells. Artificial intelligence now aids amplicon-based classification, improving speciation accuracy in mixed templates. Cutting-edge studies in 2024–2025 demonstrate the feasibility of direct-from-sample metagenomics with nanopore sequencing, providing not only species identification but also AMR gene prediction within 48 hours—a development that promises to redefine diagnostic timelines in both human and veterinary settings (10,39).
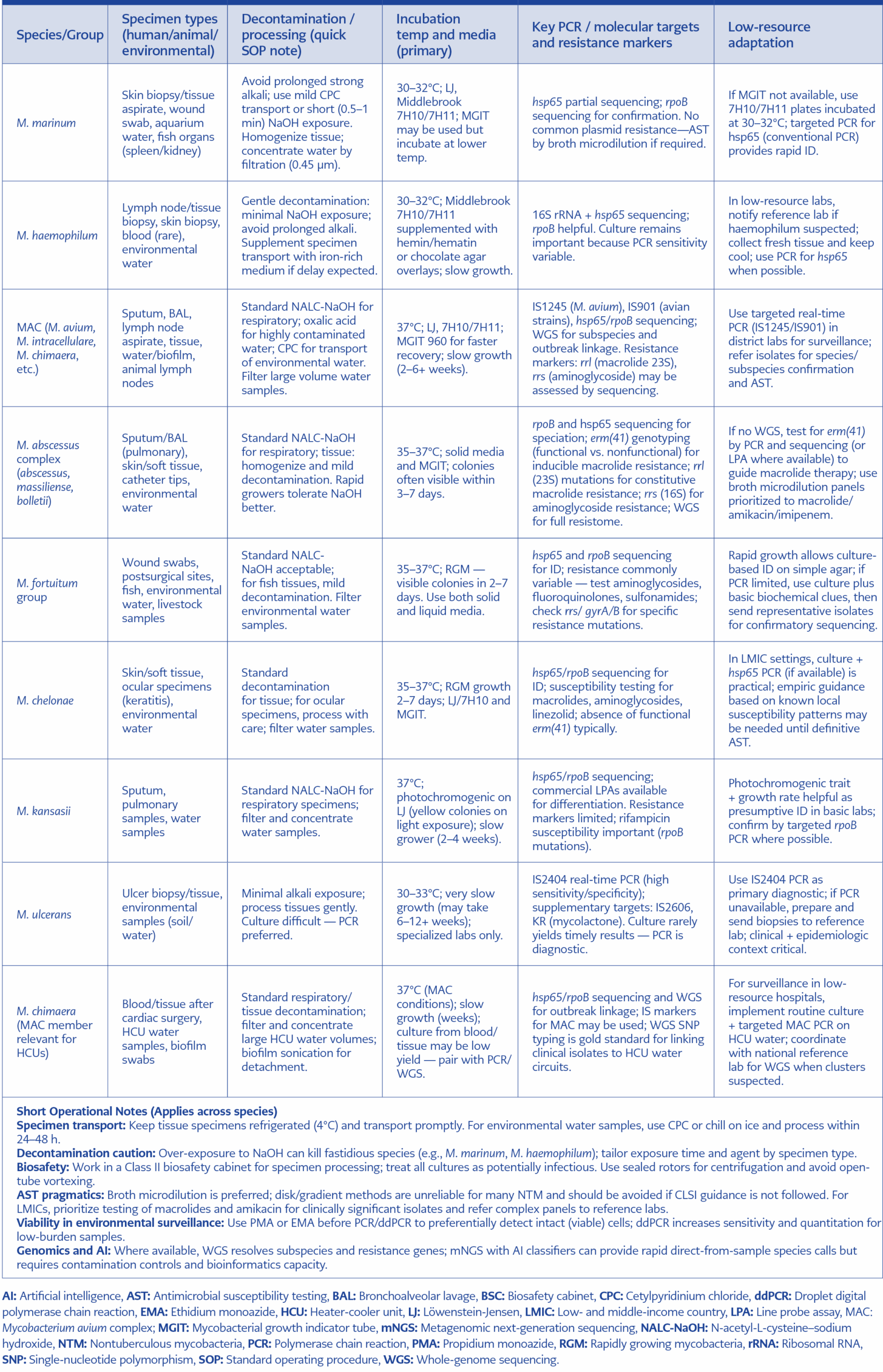
Table 4. Species-specific SOP snippets for zoonotic NTM.
Use targeted hsp65/rpoB or insertion sequence (IS)-based PCR for rapid diagnosis and PMA-ddPCR for environmental surveillance, with metagenomics reserved for unresolved or outbreak-level investigations. Species-specific SOP snippets for zoonotic NTM is summarised in Table 4.
Diagnostic Algorithms
Diagnostic approaches to zoonotic NTM have traditionally been fragmented across human and veterinary medicine, but recent frameworks emphasize a tiered, integrated strategy that leverages genomics and AI to accelerate detection and interpretation (14,40,43).
We explicitly separate clinical diagnostics (tests intended to inform patient care) from environmental surveillance (tests intended to detect and quantify organism burden in water, soil, or engineering systems). While methods overlap, the sampling strategy, reporting thresholds, and public health response differ between these use cases.
We propose a four-tier diagnostic/surveillance pathway that scales with resource availability and analytic need:
Tier 1 – Conventional microscopy and culture. Acid-fast smear and culture remain the primary initial tests for clinical specimens and for many surveillance programs; culture provides isolates for AST and WGS.
Tier 2 – Targeted molecular assays. Multiplex real-time PCR assays and targeted sequencing (hsp65, rpoB, IS elements) enable faster species-level calls and are pragmatic for LMIC reference or district laboratories.
Tier 3 – Whole-genome sequencing (WGS). WGS is the reference method for outbreak investigation, subspecies assignment, and resistome analysis when isolates are available.
Tier 4 – Metagenomics and AI-assisted pipelines (emerging). Metagenomic sequencing and machine-learning classifiers can provide direct-from-sample species calls and in silico AMR predictions in research or specialized public health centers; these approaches are promising but, as of 2025, remain largely investigational and available only at select centers.
At the first tier, conventional microscopy and culture remain the foundation in both clinical and veterinary laboratories, offering broad surveillance with modest infrastructure needs. Acid-fast bacilli smears, combined with selective decontamination and culture on solid or liquid media, provide the initial screen.
The second tier introduces targeted molecular tools. Multiplex PCR assays targeting hsp65, rpoB, or species-specific insertion sequences (e.g., IS2404 for M. ulcerans) are now widely available and suitable for LMIC settings. These assays allow laboratories to confirm NTM presence and speciate rapidly, often within 24 hours of sample receipt. In veterinary contexts, field-deployable PCR platforms have proven useful for aquaculture and livestock surveillance, enabling same-day detection of M. marinum or M. fortuitum in outbreak settings.
The third tier emphasizes high-resolution genomics. Whole-genome sequencing has emerged as the reference method for outbreak investigation, source tracking, and resistance gene profiling. In aquaculture, for instance, WGS was applied to map a 2024 M. marinum outbreak in commercial fish farms, demonstrating clonal spread across geographically separated ponds and linking fish isolates to human cutaneous infections in farm workers. Similar strategies have clarified hospital outbreaks of M. chimaera linked to HCUs, as well as zoonotic transmission chains in slaughterhouses where M. abscessus was recovered from both animal effluent and human respiratory samples.
The fourth tier integrates AI-driven diagnostics and metagenomics. Recent studies show that metagenomic next-generation sequencing (mNGS), when coupled with machine learning classifiers, can deliver species-level calls directly from raw reads within 12–24 hours, even from complex specimens such as slaughterhouse aerosols or aquaculture pond water. AI-enhanced models can also predict AMR genotypes with high concordance to phenotypic AST in recent method-development studies, offering clinicians and veterinarians actionable guidance before culture results are available (9,44–47). [Note: Reports of >90% concordance for AMR prediction and 12–24 hour turnaround times are from recent method-development studies and pilot projects; these represent emerging capabilities and should not be interpreted as widely implemented routine practice.]
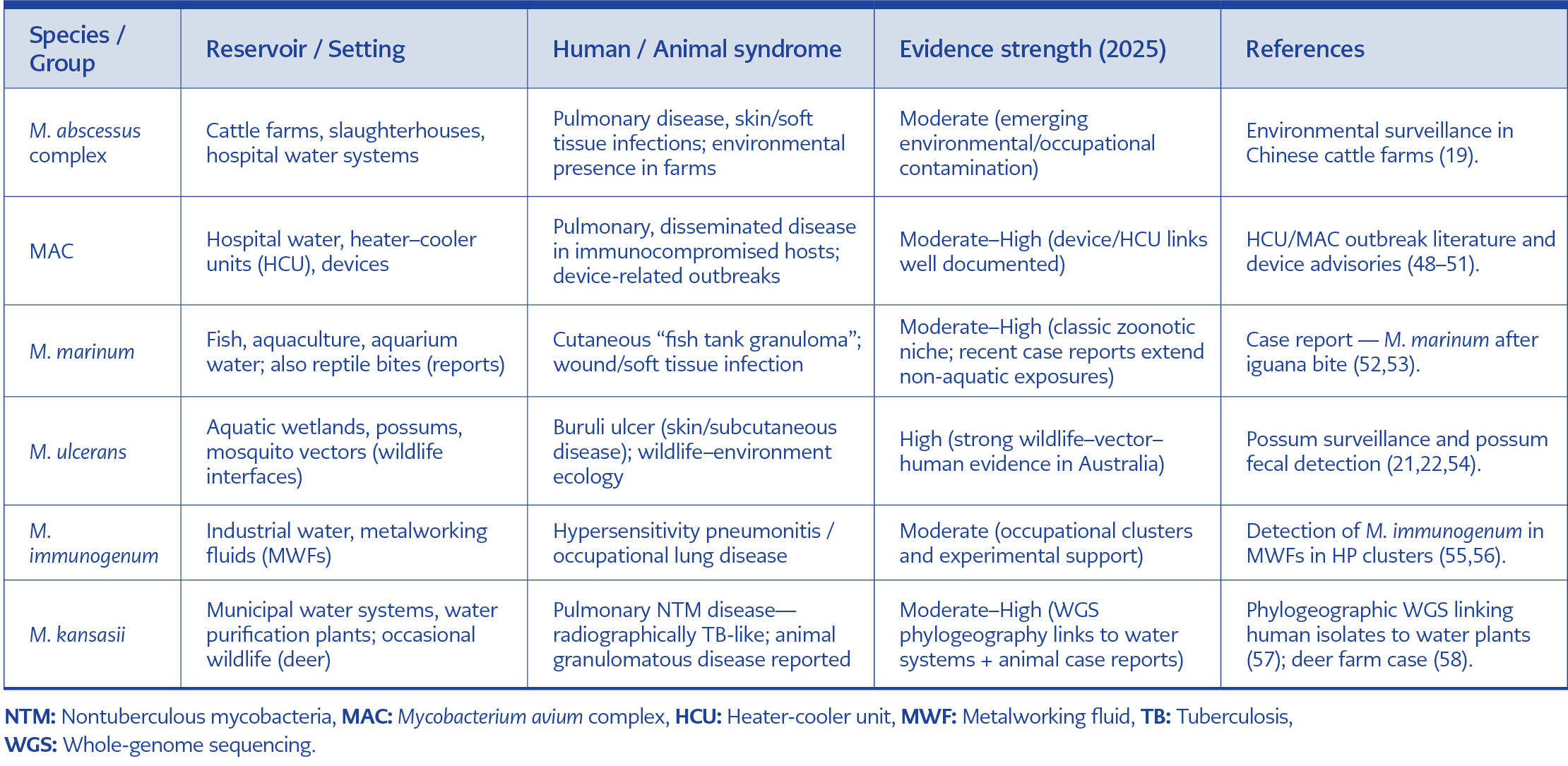
Table 5. Priority zoonotic NTM species.
Such a tiered algorithm, ranging from microscopy and culture through targeted PCR, WGS, and AI-assisted metagenomics, provides a scalable roadmap adaptable to local resources. In LMICs, targeted PCR plus selective AST can serve as a pragmatic baseline, while well-resourced national or regional centers can incorporate WGS and AI pipelines for outbreak investigation and surveillance. By explicitly linking human, veterinary, and environmental workflows, these diagnostic algorithms embody the One Health approach, bridging gaps between clinical practice, animal health, and environmental monitoring (Table 5).
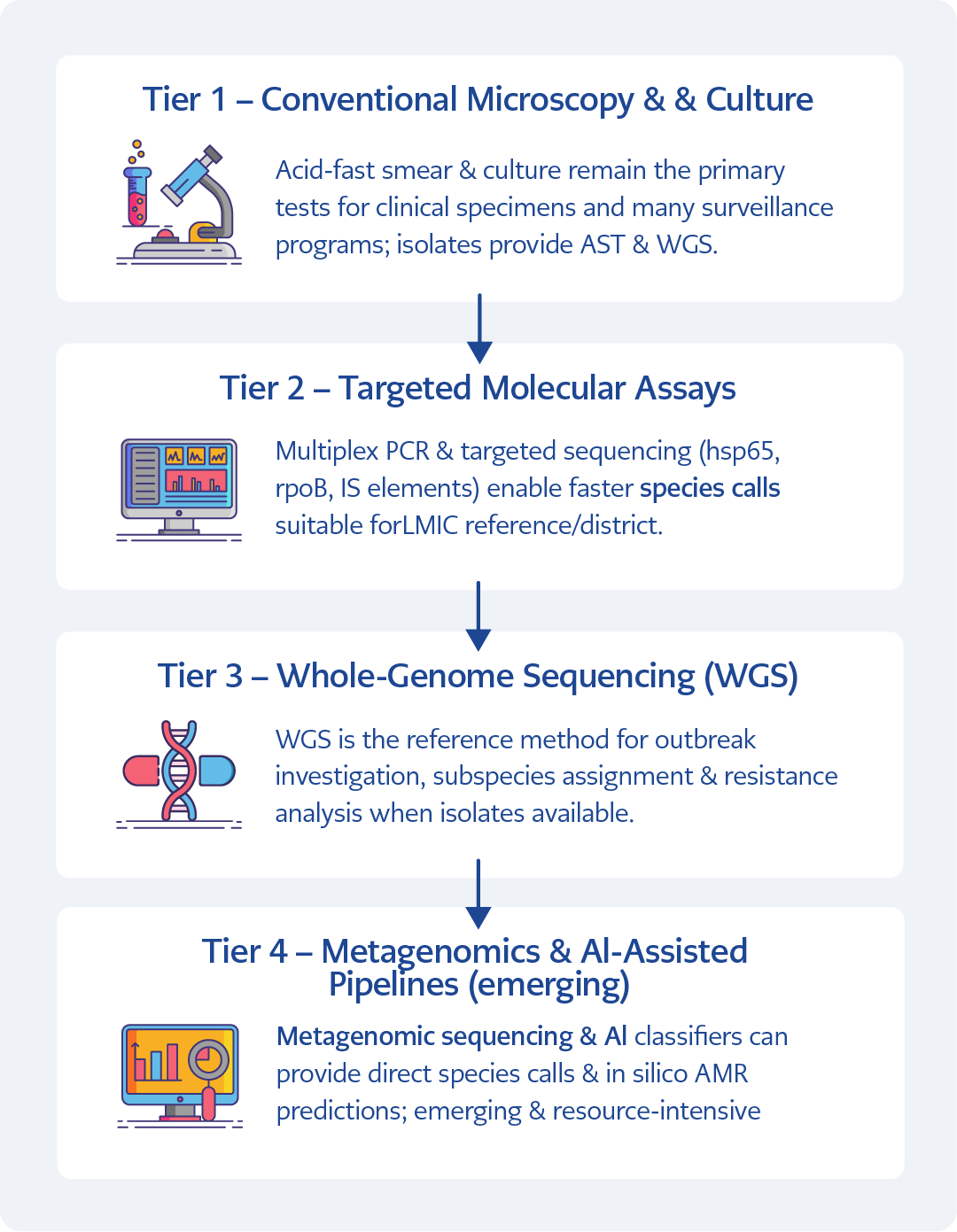
Figure 2. The figure illustrates a four-tier framework that scales from conventional microscopy and culture (Tier 1), through targeted molecular assays (Tier 2), whole-genome sequencing (Tier 3), and emerging metagenomic and AI-assisted pipelines (Tier 4). The algorithm distinguishes clinical diagnostics from environmental surveillance and shows how escalating analytical resolution supports species identification, outbreak attribution, and antimicrobial resistance prediction within a One Health context. [Graphically prepared with the assistance of ChatGPT (OpenAI, San Francisco, USA); all data interpretation and final figure content were verified by the authors.]
To accommodate the wide variation in laboratory capacity and surveillance needs across One Health settings, we propose a scalable four-tier diagnostic framework for zoonotic NTM (Figure 2).
Biosafety and Aerosol Controls
Zoonotic NTM are environmental organisms with a pronounced propensity to persist in water systems and biofilms and to become aerosolized under common engineering conditions. Recent guidance and outbreak analyses through 2024–2025 emphasize aerosol control as a central biosafety priority in both inpatient and occupational settings. Heater–cooler unit investigations continue to demonstrate that water-borne MAC members, notably M. chimaera can be aerosolized from device exhausts and contaminate operating theatres, leading to invasive infections following cardiopulmonary procedures; regulatory and clinical advisories therefore mandate strict device maintenance, water quality control, and placement/ventilation strategies to reduce aerosol dispersion (59–61). Contemporary aerosol control strategies are multi-layered and should be integrated into laboratory, hospital, aquaculture, and industrial settings where NTM exposure is plausible.
Engineered and hospital water systems as amplifiers
Hospital plumbing, endoscope washers, and HCUs can harbor NTM biofilms and serve as persistent contamination sources. Global M. chimaera outbreaks tied to HCUs illustrate how colonized devices can cause invasive disease.
Engineering controls
Source control (routine disinfection, sterile/filtered water for critical devices), closed-loop systems, correct placement and exhaust of HCUs, and enhanced building ventilation (targeted air changes per hour [ACH], high-efficiency particulate air [HEPA] filtration) are foundational mitigations. (62,63).
Administrative controls and device management
Maintain device maintenance logs, validated water-handling standard operating procedures (SOPs), pre-procedure risk assessment, and staff training. Follow device advisories and remove or decommission contaminated units as required (48,64).
Personal protective equipment (PPE) and work practices
Process aerosol-generating laboratory steps inside a certified Class II biological safety cabinet (BSC), use sealed centrifuge rotors, and employ respirators (N95 or higher) for high-load aerosol activities. In healthcare settings where device aerosol risk is identified, adopt airborne precautions for high-risk procedures (65,66).
Resource-limited adaptations
When ventilation upgrades are infeasible, prioritize single-use fluids where possible, cohort procedures to best-ventilated rooms, use portable HEPA units, and implement strict water-handling SOPs with referral mechanisms to reference labs (62,63).
Finally, biosafety planning must explicitly incorporate surveillance triggers and response actions. Thresholds for escalation should include detection of NTM in device water circuits by culture or PCR, clinical clusters of atypical postoperative infections, or repeated environmental positives despite disinfection. When triggers occur, a coordinated incident response should include device isolation, environmental remediation, patient notification and testing, and WGS-enabled linkage analysis where possible. These practices close the loop between engineering controls, laboratory detection, and clinical action — the practical embodiment of a laboratory-aware One Health biosafety program (60,61).
Reporting, Data Standards, and One Health Metadata
Rigorous reporting and interoperable metadata are essential to link human clinical cases, animal surveillance, and environmental sampling in order to clarify zoonotic pathways and inform interventions. The World Health Organization (WHO)/Quadripartite One Health agenda and regional One Health frameworks emphasize standardized data structures and multisectoral sharing to make NTM surveillance actionable. Aligning NTM reporting with these frameworks improves comparability across jurisdictions and enables rapid genomic linkage during outbreak investigations (67,68).
At minimum, every isolate or environmental sample should be reported with a standardized metadata packet that follows One Health principles: unique sample identifier; host (human/animal species or environmental); sample type and anatomical/site detail; collection date and Coordinated Universal Time (UTC) timestamp; precise geolocation (global positioning system [GPS] coordinates, with privacy controls as needed); exposure context (e.g., occupational, aquaculture, HCU-associated); specimen handling and decontamination method; laboratory methods used for detection (culture, PCR targets, mNGS/WGS) with reagent/kit versions; AST methods and breakpoints used (reference CLSI/ European Committee on Antimicrobial Susceptibility Testing [EUCAST]); and sequence accession numbers for raw reads or assembled genomes when available. For environmental and engineering-system surveillance, reporting thresholds (e.g., ddPCR copy number, metagenomic read counts, or culture colony-forming units [CFU]) should be predefined and distinct from clinical diagnostic thresholds to avoid over-interpretation of low-level colonization. These elements align with WHO and European Food Safety Authority (EFSA) guidance for integrated surveillance metadata and make downstream analyses (phylogeography, source attribution, resistome mapping) possible (69,70).
Data harmonization should also adopt controlled vocabularies and ontologies (e.g., National Center for Biotechnology Information [NCBI] BioSample fields, Minimum Information about any (x) Sequence [MIxS] environmental packages, and Open Biological and Biomedical Ontologies [OBO] Foundry ontologies) to ensure machine-readability. Where national One Health frameworks exist (for example, the United States National One Health Framework released in 2025), reporting pipelines should map local fields to these national schemas to enable federated data sharing. Metadata governance, including data ownership, privacy protections for sensitive geolocation or patient data, and standardized minimum reporting timelines, must be agreed upon by multisector stakeholders before large-scale sharing (12,70).
For genomic data, deposition of raw reads and assembled genomes to open repositories (Sequence Read Archive [SRA], European Nucleotide Archive [ENA]) with a minimal One Health metadata set accelerates outbreak response and global learning. Whole-genome sequencing-based single-nucleotide polymorphism (SNP) or core genome multilocus sequence typing (cgMLST) typing results should be accompanied by standardized quality metrics (coverage, breadth, assembly N50) and clear provenance (lab, method, date). AI- and machine-learning-derived inferences (for species identifications or AMR prediction) should be reported with model versioning, confidence scores, and training data provenance to enable reproducibility and avoid misinterpretation. Recent technical work on One Health data integration provides practical templates for linking such genomic and epidemiological data streams (69).
Operational recommendations for resource-limited settings: implement a two-tier reporting model. Tier A (local): immediate reporting of culture-confirmed or PCR-positive zoonotic NTM isolates to the local health/veterinary authority with core metadata (sample ID, date, host, specimen type, exposure setting). Tier B (reference/regional): comprehensive submission including WGS data, extended metadata, and AST details to national and international repositories when cluster signals or device-associated risks are suspected. This pragmatic model balances the need for rapid local action with centralized analytic capacity for detailed One Health interpretation (70).
One Health Surveillance and Research Agenda
Advancing understanding of zoonotic NTM requires coordinated, multinational One Health surveillance that links human clinical data, veterinary findings, and environmental sampling with genomic and exposure metadata. The VALIDATE (Vaccine development for complex intracellular neglected pathogens) network’s 2024 meeting and associated workshop reports emphasized critical gaps in integrated repositories, standardized metadata, and cross-sector pipelines that can rapidly connect an environmental or animal isolate to human clinical cases — a deficiency that directly impedes outbreak attribution and timely interventions. Building federated, interoperable repositories that accept culture isolates, raw genomic reads, and standardized One Health metadata (host, specimen type, geolocation, exposure context, and laboratory methods) is therefore a central priority (71,72).
Operational research priorities include sentinel surveillance in high-risk settings such as aquaculture, abattoirs, and hospital water systems; prospective paired sampling studies that concurrently collect human, animal, and environmental specimens during suspected clusters; validation studies for PMA-ddPCR and mNGS workflows to quantify viable NTM in water systems; and implementation science on how WGS and AI can be integrated into routine public health practice in low- and middle-income countries. These priorities reflect the consensus at recent One Health workshops that technical advances alone are insufficient without investment in data governance, workforce training, and sustainable laboratory networks (71,73).
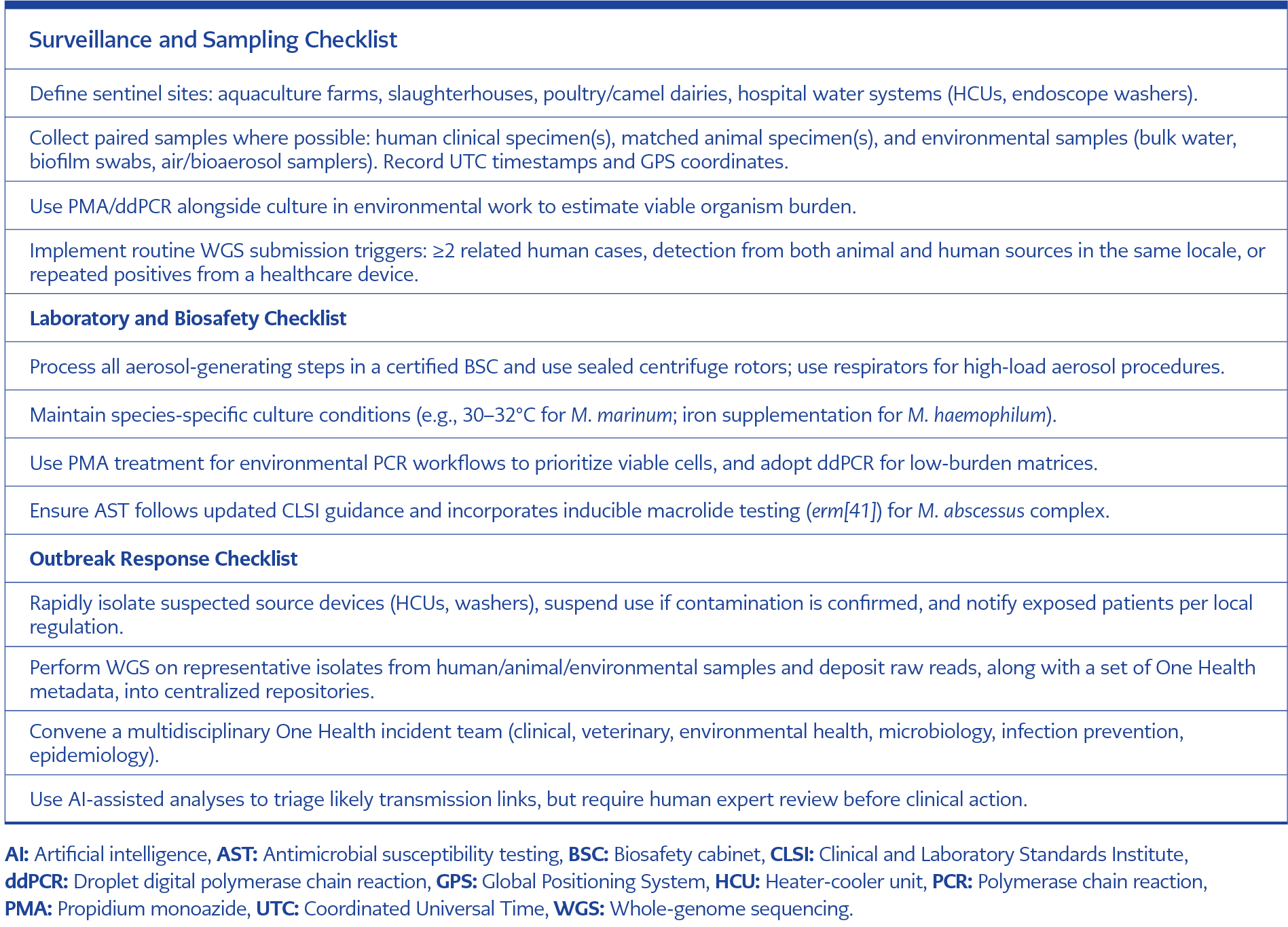
Table 6. The checklists below are operational tools intended for multidisciplinary One Health teams (clinical, veterinary, environmental, laboratory, and epidemiology) to use during routine surveillance or outbreak response.
Finally, research agendas must incorporate climate-sensitive surveillance: mapping how rising temperatures, altered hydrology, and extreme weather events reshape NTM environmental loads and human exposure patterns. This climate lens is crucial because warming aquatic systems and disrupted sanitation increase biofilm formation and persistence, expanding zoonotic interfaces in both rural and urban settings. Integrating climate data with genomic surveillance will enable predictive models that identify emergent hotspots for zoonotic NTM transmission (7,74). Much of the climate-AMR linkage for NTM is currently mechanistic or inferential; direct, pathogen-specific empirical datasets remain limited and should be prioritized in future research (Table 6).
Knowledge Gaps and Research Challenges
Despite methodological advances, several high-priority knowledge gaps limit our ability to accurately characterize zoonotic NTM risk and design interventions.
True zoonosis versus shared-source exposure
Many putative “zoonotic” events reflect shared environmental sources (for example, the same contaminated water system exposing both animals and humans) rather than direct animal-to-human transmission. Distinguishing direct zoonosis from common-source exposure requires dense sampling, finely resolved genomics (high-quality WGS with SNP-level comparisons), and robust epidemiologic linkage data. Numerous recent outbreak reports note this ambiguity and call for standardized criteria to define a bona fide zoonotic transmission event.
Integrated repositories and metadata standards
The VALIDATE 2024 reporting and workshop outputs explicitly identified the absence of interoperable repositories and agreed metadata standards as a bottleneck, making cross-study comparisons and outbreak linkage slow and error-prone. Without federated systems that allow secure sharing of sequence reads plus harmonized exposure metadata, the WGS utility for One Health attribution will remain limited. Investments in governance, legal frameworks, and finance mechanisms are essential.
Climate-AMR intersections
Climate change alters environmental conditions in ways that may both increase NTM abundance and accelerate horizontal gene transfer or selection for resistant strains. Recent scoping reviews (2023–2025) document plausible mechanisms: higher temperatures can increase bacterial replication and mobile genetic element activity; flooding and wastewater overflow can spread antibiotic residues and resistant organisms across ecological boundaries; and ecological stressors can select for both survival traits and resistance determinants. However, empirical, pathogen-specific studies for NTM remain sparse. This represents a pressing research frontier — especially in settings where warming trends and intensive agriculture co-occur (75).
Translational gaps for LMICs
Although genomic and AI tools are transformative, their operationalization in low-resource settings lags. Pragmatic validation of reduced, targeted PCR panels, PMA-ddPCR viability assays, and simplified WGS pipelines (e.g., nanopore workflows with federated cloud analysis) is needed to ensure that One Health genomics is equitable and actionable globally. Workshops and pilots should prioritize local capacity building, data governance, and sustainable financing — recommendations strongly echoed by recent One Health implementation reviews.
AMR surveillance harmonization
Veterinary and environmental AMR testing often uses different methods and interpretative criteria than those used by clinical microbiology labs. Harmonizing AST methods (or at least mapping equivalences) across sectors for NTM is essential to detect and respond to the emergence of shared resistance determinants. Global initiatives to reduce antimicrobial pollution, regulate antibiotic use in agriculture, and monitor wastewater for resistance genes will all contribute to this aim, but targeted NTM AMR studies are required to define risk and set action thresholds.
Closing Note — Pragmatic Next Steps
To translate these priorities into impact, we recommend a set of near-term deliverables: a VALIDATE-backed pilot to build a federated One-Health NTM repository with a minimum metadata standard; multisite sentinel pilots combining PMA-ddPCR environmental surveillance with paired human and animal sampling; a technical working group to define criteria that distinguish direct zoonosis from shared-source exposure; and funding mechanisms to support LMIC implementation of streamlined WGS plus AI pipelines. Together, these actions will close immediate operational gaps while building the evidence base to manage zoonotic NTM in an era of climate change and rising AMR.
Conclusion
Zoonotic NTM remain a neglected frontier at the interface of human, animal, and environmental health. Despite cataloguing more than 200 NTM species, only a fraction has been interrogated for their zoonotic potential, and critical questions persist over whether observed cross-species infections represent true transmission events or shared environmental exposures. The last five years have revealed alarming patterns: rising M. abscessus pulmonary disease epidemiology and detection of Mycobacterium immunogenum in industrial water systems. These shifts are compounded by accelerating climate change, which expands aquatic and soil reservoirs, and by AMR trends that complicate therapy in both human and veterinary medicine.
Diagnostic innovation ranging from low-resource PCR assays to AI-enabled metagenomics provides a framework for detection, but translation into harmonized One Health surveillance is lagging. Equally pressing are biosafety measures: recent 2025 guidance highlights the need for strengthened aerosol control in laboratories, while reporting standards must now align with WHO One Health metadata frameworks to ensure interoperability.
What is urgently needed is a coordinated global investment in integrated surveillance systems that leverage genomics, veterinary fieldwork, and environmental microbiology, while embedding NTM into broader AMR and climate-health agendas. Without such action, the zoonotic dimensions of NTM will remain underestimated, and opportunities to mitigate outbreaks in farms, clinics, and communities will be lost. We call upon funders, policymakers, and research consortia to treat zoonotic NTM as a priority pathogen complex and to support collaborative platforms that unite molecular epidemiology, One Health reporting, and cross-sectoral training. Only with such integrated vigilance can we anticipate, rather than merely react to, the next wave of NTM threats.