Introduction
The straightforward way to keep the course of the pandemic under control is herd immunity, which is established with a vaccine against severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). There is a need to develop appropriate laboratory tools to predict the efficacy of vaccines and evaluate the post-vaccination immune response (1). However, several studies have shown that circulating antibodies gradually decrease in patients recovering from coronavirus disease 2019 (COVID-19) (2, 3). These findings triggered boosting doses of COVID-19 vaccines. As of May 22, 2023, a total of 13,355,264,024 vaccine doses have been administered worldwide. The number of persons vaccinated with at least one dose is 5,550,147,252; fully vaccinated with the last dose of primary series is 5,108,289,510 (4). Although booster doses were administered widely, the best schedule is not clear for all vaccine types, particularly in countries where both inactivated virus vaccines and mRNA-based vaccines are available.
The SARS-CoV-2 vaccination program in Türkiye started with the inactivated virus vaccine CoronaVac (Sinovac Biotech Ltd., China) in January 2021. Then, mRNA vaccine BNT162b2 (Pfizer-BioNTech, Germany) was added to the vaccination schedules. The emergence of the Omicron variant, which was first reported in November 2021 and quickly became globally dominant, was associated with the potential for immune escape from vaccine-induced antibodies (5). In Türkiye, a policy of boosting with CoronaVac or BNT162b2 allowed for evaluating the effect of homologous and heterologous boosting in individuals primed with an inactivated vaccine.
Recent research has highlighted that robust and durable T-cell immunity may be a more reliable marker of immunity against COVID-19 (6, 7). Therefore, it may be important to measure both antibody production and T-cell response when assessing the strength and duration of the immune response to SARS-CoV-2 infection and vaccination in patient groups. Interferon-gamma (IFN-γ) release assays (IGRAs) are the most frequently used methods for the evaluation of T-cell response following infection and/or vaccination (8–12).
This study aimed to determine the effects of CoronaVac and BNT162b2 boosters on the humoral and cellular immunity of individuals primed with two doses of CoronaVac.
Materials and Methods
Study Design and Participants
The study, which is a prospective, single-center, and observational cohort study, was conducted at Hacettepe University Hospital, Türkiye, between February 2022 and December 2022. Serum samples of participants aged 18–59 years, without any comorbidities, immune-mediated systemic diseases, or autoimmune diseases, and who were not under immunosuppressive drug therapy and/or disease-modifying antirheumatic drugs were included in the study. Participants with no clinical history of COVID-19 who were immunized with three doses of CoronaVac (Group 1), two doses of CoronaVac and one dose of BNT162b2 (Group 2), and two doses of CoronaVac plus two doses of BNT162b2 (Group 3) were included in the study. The humoral and cellular immune responses of the participants were assessed within 1–12 months after the last dose of vaccination. Sampling time was taken into consideration, as it varied for each patient.
Blood samples were obtained both for immunoglobulin G (IgG) assay and for IGRA only at the first visit. For the IGRA, lithium heparin whole blood samples were used. Demographic characteristics of the individuals were recorded. All participants were followed up to six months after sampling, and any SARS-CoV-2 infection confirmed by a positive PCR test was noted during the follow-up period. No antibody and IFN-γ assays were repeated during the follow-up. Serum samples for IgG were stored at -80ºC until assayed.
All experiments were carried out in compliance with relevant laws and guidelines and with the ethical standards of the Declaration of Helsinki. The study was approved by the Hacettepe University Clinical Research Ethics Board on March 15, 2022, with decision no 2022/02-05 and KA-21156, and by the Non-Interventional Clinical Research Ethics Board on March 16, 2021, with decision no 2021/06-45 and GO 21/366. Written informed consent was obtained from all participants prior to enrolment.
Immunological Assays
Detection of SARS-CoV-2 antibodies
Quantitative IgG levels against the spike receptor binding domain (RBD) protein of SARS-CoV-2 were tested with the chemiluminescent microparticle immunoassay method using the SARS-CoV-2 IgG II Quant assay (Abbott Diagnostics, USA) on the Architect i2000SR analyzer (Abbott Diagnostics, USA). Antibody concentrations obtained in AU/mL were multiplied by the correlation coefficient of 0.142 and expressed as binding antibody units (BAU/mL) according to the World Health Organization (WHO) International Standard for anti-SARS-CoV-2 immunoglobulins (13). Concentrations of 50 AU/mL (7.1 BAU/mL) and above were considered positive. This test has been reported to be 100% compatible with the plaque reduction neutralization test (PRNT), and a concentration of 149.1 BAU/mL was associated with a 1:80 dilution of PRNT (14).
SARS-CoV-2 interferon-gamma release assay (IGRA)
IFN-γ release from SARS-CoV-2 specific T cells was determined quantitatively by the Quan-T-Cell SARS-CoV-2 and Quan-T-Cell ELISA kits (EUROIMMUN, Germany) according to the manufacturer’s instructions. Briefly, 500 μL of whole blood from lithium-heparin blood collection tubes was pipetted into three separate stimulation tubes and incubated at 37°C for 20–24 h.
The stimulation tubes used were as follows: 1) IGRA BLANK, without IFN-γ activating substance for the determination of the individual IFN-γ background; 2) IGRA TUBE, containing antigens based on the S1 domain of the SARS-CoV-2 spike protein; and 3) IGRA STIM, containing a mitogen for non-specific T-cell stimulation for control of the viability and stimulation ability. After the incubation, the tubes were centrifuged at 12,000 g for 10 min, and 200 μL plasma was stored at 2–8°C until testing. Plasma was analyzed for IFN-γ levels by the IFN-γ ELISA kit (EUROIMMUN, Germany). IFN-γ concentration was expressed as mIU/mL. A sample result was considered valid when IFN-γ values were <400 mIU/mL in IGRA BLANK and >400 mIU/mL in IGRA STIM – IGRA BLANK. Samples with IFN-γ values above the standard curve were further diluted 1:20, depending on the O.D. of the initial measurement. The results were interpreted according to the manufacturer’s recommendations: IFN-γ <100 mIU/mL as negative, 100–200 mIU/mL as borderline, and >200 mIU/mL as positive.
Statistical Analysis
Statistical analyses were performed using IBM SPSS Statistics software, version 23.0 (IBM Corp., USA). The Shapiro-Wilk normality test was used to assess the normality of the distribution of numerical variables. The Kruskal-Wallis test was used to compare numerical variables across more than two independent groups. Dunn’s post hoc test was applied for the significant results of the Kruskal-Wallis test. Pearson’s chi-square analysis was used to determine whether there was a significant relationship between categorical variables. The difference between the two independent groups in terms of numerical variables was evaluated with the Mann-Whitney U test. The statistical significance was set as p<0.05.
Results
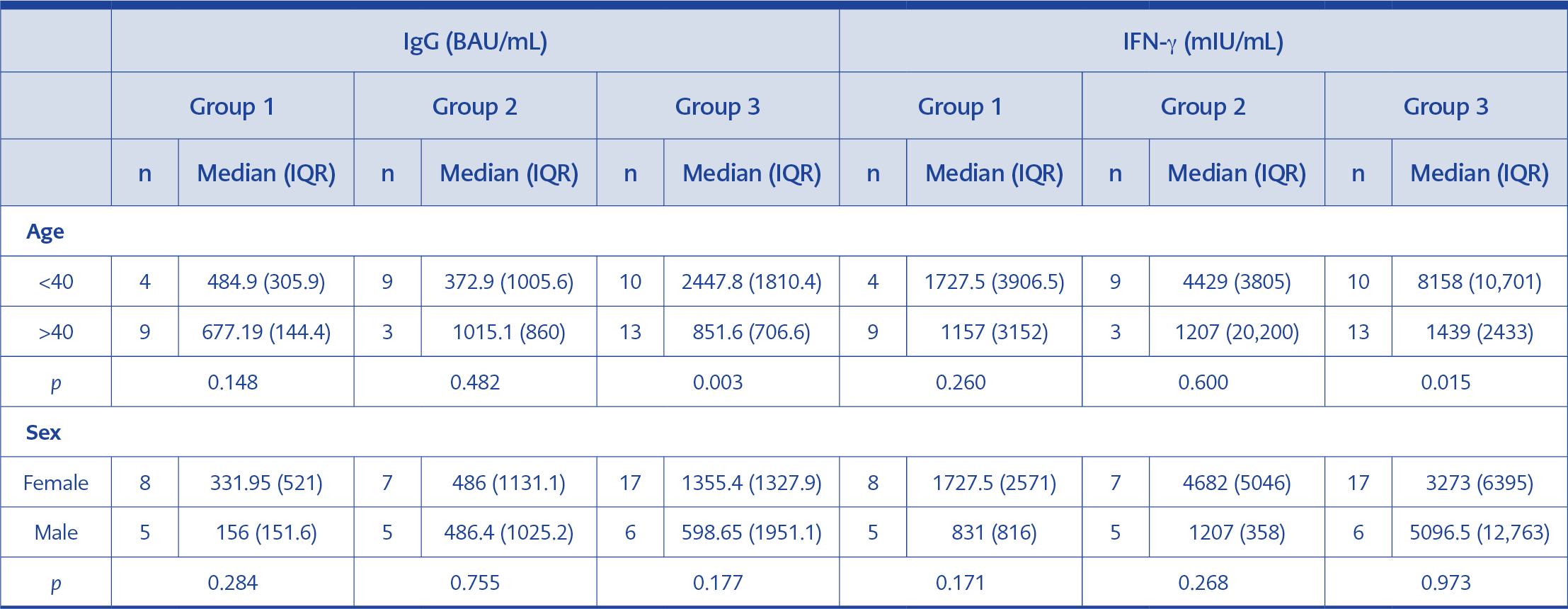
Table 1. Comparison of IgG and IFN-γ values by gender and age for the study groups.
Serum samples of 48 participants were included in the study. The age and gender distribution of the individuals in the study groups is presented in Table 1. No significant gender difference was found for IgG and IFN-γ values across the three groups. In Group 3, the participants under the age of 40 had significantly higher IgG and IFN-γ median values than those over 40 (p=0.003 and p=0.015, respectively). No significant difference was found in the other two groups.
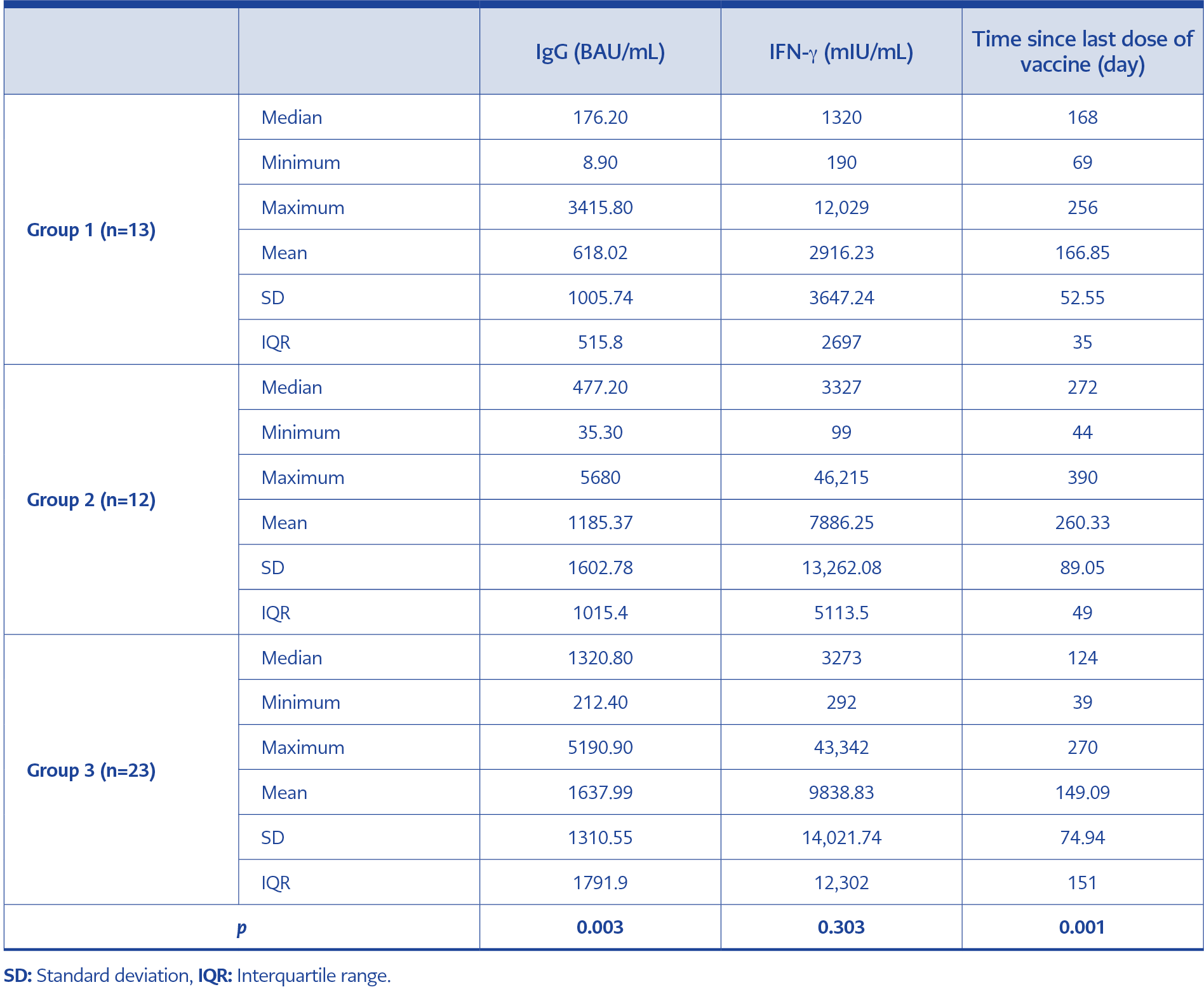
Table 2. Quantitative IgG and IFN-γ levels in the study groups and time after the last vaccination date.
SARS-CoV-2 IgG seropositivity was detected in all participants across the study groups. The median IgG values for Group 1, Group 2, and Group 3 were 176.20, 477.20, and 1320.80 BAU/mL, respectively. The median value of the IgG levels determined in the three groups was above the concentration of 149.1 BAU/mL, which was associated with a 1:80 dilution of PRNT. Quantitative IgG analysis revealed a significant difference between Group 1 and Group 3 (p=0.003). The highest IgG levels were detected in individuals boosted with two doses of BNT162b2 (Group 3). The median IgG level of Group 2 was not significantly different from the other groups (Table 2).
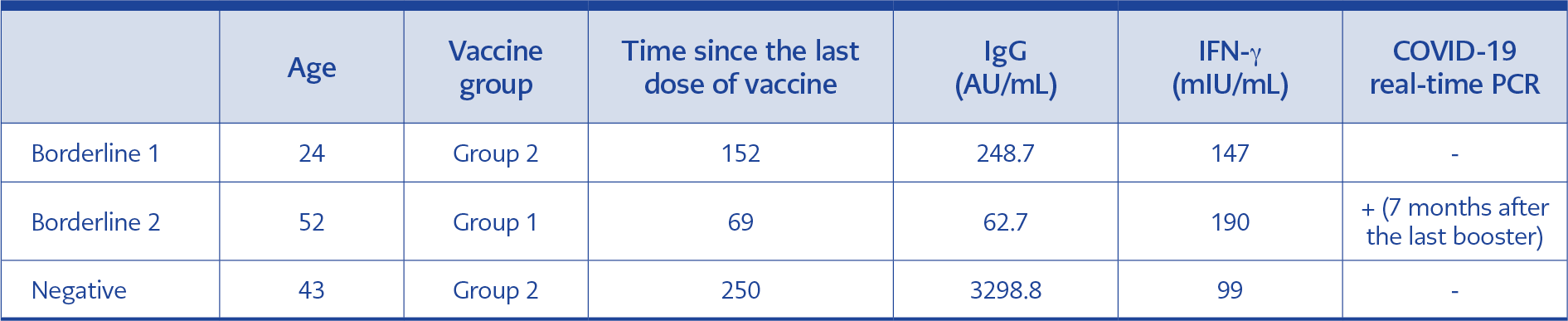
Table 3. Characteristics of participants with borderline and negative IFN-γ results.
The highest levels of IFN-γ response were observed in Group 3, while the lowest were seen in Group 1. However, there was no significant difference among the three groups regarding IFN-γ values (p=0.303). The median IFN-γ values of Group 1, Group 2, and Group 3 were 1320.0, 3327.0, and 3273.0 mIU/mL, respectively (Table 2). There were two borderline and one negative IFN-γ result among the participants, whose details are given in Table 3.
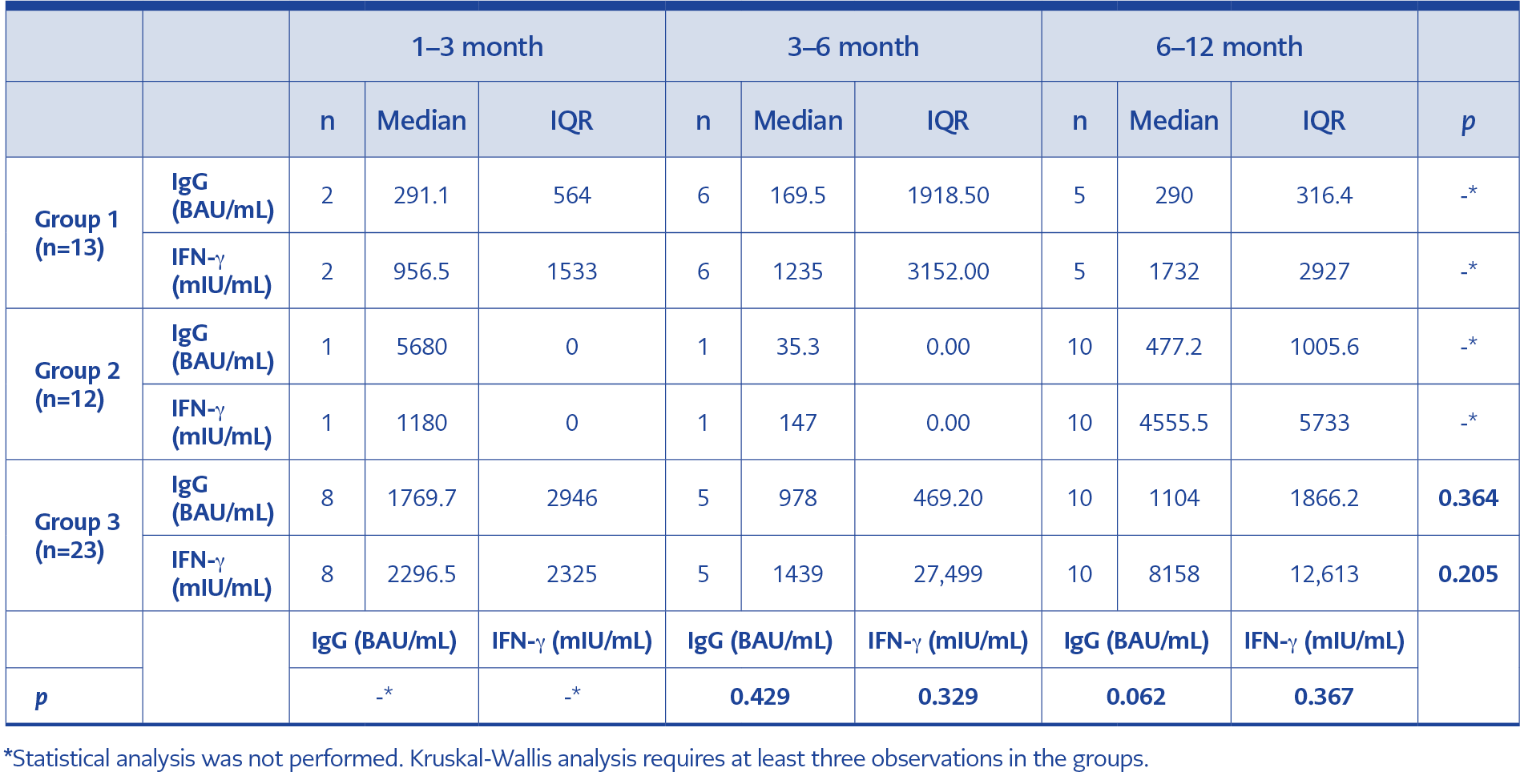
Table 4. Values of intergroup parameters in terms of elapsed time following the last booster.
Sampling time since the last vaccine dose varied among the groups. The difference between the groups in terms of sampling time was significant (p=0.001) (Table 2). To evaluate the impact of time passed since the last vaccination, IgG and IFN-γ values of the three groups were analyzed in terms of elapsed time in three monthly periods (Table 4). There was no significant difference in IgG and IFN-γ values (p=0.364, p=0.205, respectively) regarding the time elapsed for Group 3. Statistical analysis could not be performed for Group 1 and Group 2 since the number of observations was less than three for some time intervals (Table 4).
No statistically significant results were found for IgG and IFN-γ among the three groups that were compared in the 3–6 month and 6–12-month periods. Group 2 was excluded from the 3–6 month analysis because this group had only one observation. The number of observations in Group 1 and Group 2 was insufficient for statistical analysis in the 1–3-month period (Table 4).
The participants were followed up until December 2022. During the follow-up period, 23 (47.92%) of the 48 participants tested positive for COVID-19 by SARS-CoV-2 real-time PCR. The PCR positivity during the follow-up period was 8/13 (61.54%) in Group 1, 4/12 (33.33%) in Group 2, and 11/23 (47.83%) in Group 3. None of the participants who had COVID-19 were hospitalized. A statistically significant relationship was found between COVID-19 status and age (p<0.001). The rate of positive COVID-19 results in participants over the age of 40 (72%) was significantly higher compared to those under 40 (21.74%). The median time interval between the administration of the last vaccination and the onset of COVID-19 was 223 days. The median time interval between the immunological investigations to ascertain vaccine response and the subsequent COVID-19 infection was 100 days. There was no statistically significant relationship between COVID-19 status and gender, vaccination status, or the time elapsed monthly from sampling (p>0.05).
Discussion
Heterologous prime-boost vaccine regimens are being discussed globally to close the COVID-19 vaccine supply gap and improve the effectiveness of immunogenicity. Heterologous prime-boost vaccination is based on a strategy mixing different vaccine types to present the same antigen to the immune system in a different way so that a stronger immune response is provided. This study aimed to compare the humoral and cellular immune responses after homologous and heterologous COVID-19 boosters in CoronaVac-primed individuals.
The data available for comparing humoral and cellular immunogenicity achieved using CoronaVac or BNT162b2 as the booster in individuals following priming with the CoronaVac are limited. Our study is important for providing data in that respect. The humoral response to SARS-CoV-2 is strongly associated with protection, and the T-cell response, which indicates cellular immunity to SARS-CoV-2, is considered important in preventing severe clinical outcomes (1). As new variants of SARS-CoV-2 have shown the ability to evade vaccine-induced neutralizing antibodies, the role of T-cell immunity has been increasingly explored (12, 15, 16).
This study provides real-life data on IgG levels following homologous CoronaVac and heterologous mRNA booster doses at various time points after priming with two doses of the inactivated CoronaVac vaccine. In the 6–12-month post-vaccination period, the lowest quantitative IgG levels were detected in Group 1, which received a CoronaVac booster. IgG levels were higher in the two groups that received BNT162b2 boosters, with Group 3 (two BNT162b2 boosters) showing the highest levels. This finding was also reflected in the higher number of participants who developed COVID-19 following the last booster in Group 1. Thus, booster doses with CoronaVac may raise a concern regarding waning humoral immunity and decreasing protection, particularly against SARS-CoV-2 variants. However, IgG BAU/mL levels were still at a considerable amount in the 6–12-month period in Group 1 with the homologous application of three doses of CoronaVac. This result also provides data on the duration of humoral immunity in individuals who received only inactivated vaccines. Although the rate of COVID-19 development was highest in that group, it was noteworthy that none of the RT-PCR-positive cases required hospitalization.
The decrease in median IgG and IFN-γ results from the 1–3-month period to the 3–6-month period, as well as the increase from the 3–6-month period to the 6–12-month period (Table 4), should not be interpreted as a decrease or an increase in immune response, as the participants were not followed up longitudinally. Participants followed in each period were different individuals; in Group 3, the number of participants was 8 in the 1–3-month period, 5 in the 3–6-month period, and 10 in the 6–12-month period. These numbers were insufficient to represent a population; therefore, it can be argued that the result is coincidental.
Several studies and vaccine trials have shown that the overall effectiveness of BNT162b2 and CoronaVac is affected by many factors, including age, body mass index (BMI), smoking status, comorbidities, time post-vaccination, previous COVID-19 infection, and circulating SARS-CoV-2 variants (17). A strength of the present study is that it included participants with no known history of COVID-19 and no immunosuppression or comorbidities. Thus, some possible factors associated with a reduced immune response or complete unresponsiveness to COVID-19 vaccination were excluded. In our study, we observed that the IFN-γ response in participants over the age of 40 was lower compared to those under 40. It is known that cellular immune response decreases with age. For example, Schwarz et al. (17) reported that the IFN-γ response of healthcare workers with a mean age of 34 years who received the BNT162b2 vaccine was significantly higher compared to the elderly group with a mean age of 81 years. While some COVID-19 vaccine studies have shown that the immune response is higher in women, some studies have observed no gender difference (18). In our study, we found no difference in either cellular or humoral immune responses by gender.
Previous studies have shown that following priming with two doses of CoronaVac vaccination, a heterologous mRNA booster is more immunogenic than a homologous booster (15,19–25). An extensive cohort study performed by Sonmezer et al. (19) revealed that a third dose of the CoronaVac booster prevented 58.24% and BNT162b2 87.27% of symptomatic COVID-19 cases in the study cohort, with no hospitalizations, intensive care unit admissions, or deaths in third-dose booster groups. Kuloğlu et al. (20) found a significantly lower neutralizing antibody titer in the CoronaVac booster group than in the BNT162b2 booster group. Demirhindi et al. (21) found that the heterologous vaccination strategy “CoronaVac and BNT162b2 regimen” was able to induce a stronger humoral immune response. However, their study did not evaluate the effect of a second BNT162b2 booster. Saltoğlu et al. (22) also reported that the administration of the mRNA vaccine as a booster dose could provide more effective protection against COVID-19 infection. The results of our study supported the data that heterologous boosting should be considered to establish better humoral immunity obtained with COVID-19 vaccines.
The coordination of humoral and cellular responses is critical for an intact immune response against infections. Cell-mediated immunity is important both to prevent the development of infection and protection from severe COVID-19 disease (26). Although data about the effect of heterologous and homologous boosters on humoral immunity in CoronaVac-primed individuals are plenty in the literature, data on T-cell immunity following the homologous and heterologous boosters are still lacking. The current study aimed to provide data on the level of IFN-γ release from SARS-CoV-2 activated T cells in different booster regimens. Assessment of the T-cell response was performed in our study by the determination of IFN-γ secretion from effector T cells. IGRAs are easy to perform, and their validity to define vaccine responsiveness was shown in some standard COVID-19 cohorts (27, 28).
We observed the highest IFN-γ response in the group that received two BNT162b2 booster doses. Although the difference in the IFN-γ levels among the three groups was not statistically significant, the individuals who were boosted with the mRNA vaccine revealed two- and three-fold higher levels than the homologous boosted individuals. Whether this result may be related to the shorter time elapsed after the last vaccine dose in the two BNT162b2 boosted group is unknown. One of the two individuals who revealed a borderline IFN-γ response had a positive COVID-19 PCR result seven months after the last booster. That individual was in the homologous CoronaVac group and had the lowest level of SARS-CoV-2 specific IgG in the whole study groups. One IFN-γ borderline and one negative individual who was PCR-negative had higher specific IgG levels. Although the number of IFN-γ borderline or negative individuals was very small, these findings might indicate the more substantial effect of virus-specific IgG in protection from COVID-19 disease.
It is well known that inactivated vaccines exhibit no or minimal effect on cellular immunity (26). CoronaVac, an inactivated virus vaccine, has been shown to exert cellular immunogenicity. An increase in the IFN-γ response following CoronaVac vaccination is a part of the cellular immune response, as shown in several studies (24, 29). However, boosters with an mRNA vaccine usually lead to a more significant IFN-γ response. In the study by Lai et al. (24), the IFN-γ response in participants who received CoronaVac as a booster after two doses of CoronaVac was found to be significantly lower than those with BNT162b2 applied as the third booster. Similarly, in the study by Kanokudom et al. (25), following two doses of CoronaVac, a significant increase in IFN-γ response was observed in vector and mRNA vaccine applications as the third booster compared to a third booster application of inactivated vaccines. Kuloğlu et al. (20) concluded that the ratio of the effector T cells increased along with greater IFN-γ activation after the BNT booster following CoronaVac priming.
Inactivated virus vaccines, unlike mRNA-based vaccines, induce a T-cell response that recognizes multiple different proteins but is mediated exclusively by CD4+ T cells (30). A recent study with the apparent efficacy of inactivated-virus-based vaccines in reducing disease severity in the recent Omicron infection wave in Hong Kong suggests that the breadth of vaccine-induced multiprotein-specific CD4+ T-cell response might compensate for the absence of CD8+ T-cell responses in controlling Omicron-induced pathology (31).
One of the main limitations of this study was that the immune response of the participants was not followed up longitudinally on a monthly or yearly basis. Another limitation was the low number of participants, which might be reflected in the lack of statistical significance in spesific comparisons between the groups. However, the rapid changes in the pandemic conditions, the changing trends in vaccination regimens, and the difficulty in accessing participants who had not previously had COVID-19 were the disadvantages that inhibited the inclusion of more participants in the study. Additionally, the COVID-19 history of the participants before they were included in the study was based on their personal declaration and clinical history without additional molecular or serological testing. A further limitation was the statistically significant difference between the groups in terms of the time since the last dose of the vaccine. This prevented the evaluation of the correlation between time since the last dose of vaccination and humoral and cellular immunity levels. Although IgG antibody levels were measured using an assay that was reported to be 100% compatible with the PRNT, neither pseudovirus nor live-virus neutralization assays were performed in this study.
We conclude that although both homologous and heterologous boosting provided effective humoral and cellular immunity in inactivated vaccine-primed individuals, boosting with two doses of BNT162b2 should be prioritized, particularly in people with a higher risk of infection with SARS-CoV-2 since it exhibited a clearly positive impact on both types of immunity.